| Main authors: | Bautista, S., Urghege, A.M., Camacho, A., Turrión, D., Jaime, L., Vera, M.A., Nazarova, V., Vega-Rosete, S., Sáez-Cases, A., Fuster, A., Morcillo, L., López-Poma, R., Valera, M., D. Fuentes, and Rodríguez, F., Bladé, C. and Mayor, A.G. |
| Editor: | Jane Brandt |
| Source document: | Bautista, S. et al. (2017) Dryland restoration dynamics and thresholds as a function of plant pattern and diversity. CASCADE Project Deliverable 4.3 31 pp |
Patch size and biotic structure (composition/diversity) modulated the functioning (productivity) of the dryland communities artificially established on bare degraded soils in the framework of the manipulative restoration experiments.
For experiment #1 (Figure 1), Atriplex halimus (a pioneer, fast-growth species that easily colonizes bare-soil areas) small-patch plots produced significantly higher biomass than plots with big patches. This result was partly unexpected, as previous work in CASCADE proved that big patches are more efficient in capturing water and other resources from runoff (see »Potential for sudden shifts in ecosystems), and probably reflects an adverse effect of the competition for space in big (dense) patches that counterbalanced and exceeded the positive effect of higher runon inputs. For the grass species Lygeum spartum, this effect of patch size was marginal. Biomass production for the tall-shrub, late-successional species Phyllirea angustifolia was very low and did not show any effect of patch size (Figure 1, left). Biomass production in diverse patches (polycultures) did not significantly vary with patch size. Plot biomass from polycultures was marginally (no significant) higher than the average of the three monocultures (Figure 1, right). However, this difference was only apparent for big-patch pattern and biomass production in polycultures was similar to plot biomass from the most productive monoculture (A. halimus), which indicates that enhanced production in polycultures was not due to any functional complementarity among the species but mostly to the presence of the most productive one in the mix (Loreau and Hector, 2001).
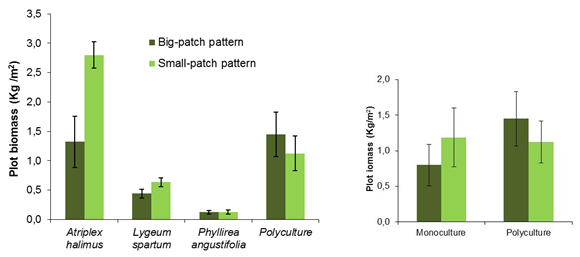
Figure 1
At early stages after planting, with all plant seedlings sharing similar rooting space, there was no sign of complementarity between species that lead to higher productivity in diverse patches (polycultures). However, there was no sign of detrimental effects of interspecific competition either, as compared with intraspecific competition in monocultures. It seemed that big-patch polycultures benefited better from the higher capacity for trapping water and other resources from runoff (see »Potential for sudden shifts in ecosystems) than big-patch monocultures, which would explain that the slight positive effect of patch diversity on the overall patch productivity was only observed for big patches.
By looking at the individual plant response (individual plant growth rate) within the different types of patches, we found that the effect of patch size and diversity on plant growth largely varied as a function of the species considered and the environmental (climatic) conditions. Figure 2 shows the results for the most contrasting species A. halimus and P. angustifolia, with results or L. spartum (data not shown) being intermediate between those from these two species.
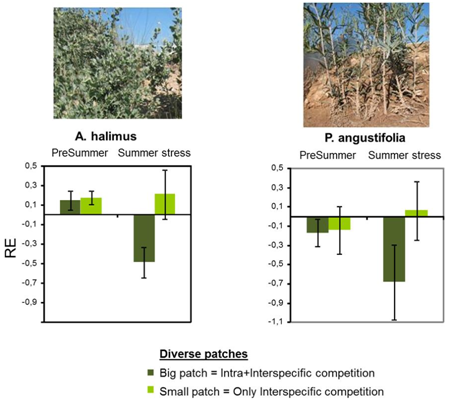
Figure 2
Under good environmental conditions (pre-summer), the pioneer, fast colonizer A. halimus performed better in polycultures, indicating that intraspecific competition in monocultures was stronger than interspecific competition in polycultures. This response was similar for big and small patches. Conversely, the slow-growth, late-successional species P. angustifolia performed better in monocultures, particularly for big patches, reflecting the adverse effect of competition with the other two species. However, under stressful conditions (summer period) both species performed better in monocultures when growing in big patches, particularly P. angustifolia.
Since big patches in polycultures included several individuals of each species, these results suggest that dealing with both intra-specific and interspecific competition under stressful condition is more challenging than interacting only with conspecific individuals. This kind of response has not been previously reported and has direct implications for restoration: in case of pursuing the restoration of multi-specific patches, it would be advisable to introduce only one individual per species, particularly if stressful conditions are expected to occur during the early stages of the restoration process.
Results from experiment#2 at the patch scale showed a significant effect of patch size for B. retusum growth rate, but not for R. lycioides growth rate. The effect of patch type (mono vs. bi-culture) was not significant in any case (Figure 3). These results are consistent with those from experiment#1 in showing (1) similar performance of both species in polycultures and bi-cultures for small patches, (2) better performance of the fast-growing species (in this case: B. retusum) in small patches, and (3) a trend towards worse performance for the late successional species (R. lycioides) when coexisting with other species in big patches (Figure 3).
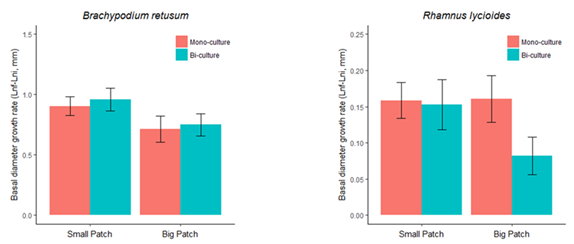
Figure 3
It is generally assumed that grasses exert strong competition over seedlings and young saplings of woody species (Gómez-Aparicio, 2009), particularly if shrub seedlings grow within a matrix of grass cover and the associated dense belowground network of grass roots.
Under a context of dryland restoration of patchy landscapes, patches that include grasses and shrubs may not be that detrimental for the shrub species, particularly if enough bare-soil interpatch area is left around the patches. Regarding the potential effect of initial cover on restoration success we did not identify any limitation to the restoration potential in the case of establishing plant communities with low, or very low, initial cover values (Figure 4). In fact, initial plant cover hardly affected plant growth, with only R. lycioides in biculture patches showing a marginally significant decrease in growth rate with plant cover.
These results challenge the conceptual framework provided by the fold-bifurcation type of models, which generally assume that dryland vegetation recovery would be very limited if starting with degraded low-cover, unless conditions largely improve. Restoration aims to act as a shortcut in the recovery dynamics of degraded drylands, facilitating the return to healthy states by manipulating either the environmental conditions or the initial state of the biotic community or both (Sudding et al., 2004). The fact that in restored patches, plants are introduced at the seedling stage may be an advantage –as compare with natural colonization from seeds– that contributes to overcome part of the limitations for recovery.
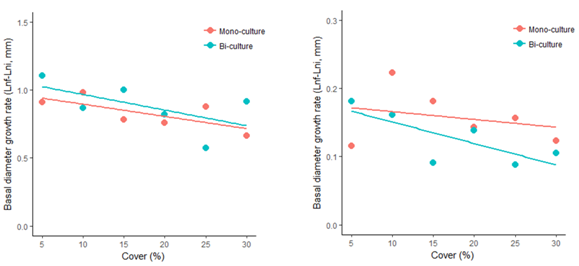
Figure 4
Given the slow growth rate of most perennial species in dryland communities, the assessment of the recovery dynamics of experimental ecosystems is limited by the relatively short duration of any single research project. Therefore, in order to address the research questions on recovery dynamics, we relied on a modelling approach (see »The role of ecohydrological feedbacks in dryland degradation reversal). However, the results from the manipulative experiments described above suggest that a low initial plant cover does not constrain the potential for restoration success, which could be explained by the positive effect of water and sediment transfer from large bare soil areas to few existent plant patches.
Finally, the large-scale experiment conducted at ES56 followed a different approach to the ES24 experiments. Instead of comparing mono and polycultures, the experiment ES56 assessed the effect of increasing patch size (number of individuals) and diversity (number of species and/or functional groups), using the performance of patches with one individual and one species as reference. Figures 5 to 7 summarize the main results obtained at the early stages of the restoration trajectory.
We found that overall survival was not significantly hampered by increasing the number of individuals and/or species (up to 8 in both cases) in the restored vegetation patch (Figure 5). However, not all plant functional groups responded in the same way to patch size and diversity (Figure 6). Survival of grasses was hardly affected by including in the patch other species and/or individuals. Conversely, survival of woody species significantly varied between treatments, decreasing from patches with only one individual (treatment 1_1) to patches with two individuals (one per species) of two species from the same functional group (treatment 2_2); and further decreasing in patches with two individuals per species of two species of the same functional group (treatment 4_2). The exception to this trend was the group of resprouter phanerophytes (tall, late successional shrubs), which showed a very high survival for treatment 4_2, probably due to the influence of one of the experimental plots of this kind that was, incidentally, in particular good conditions. Further increase in number of species and functional groups per patch (treatments 4_4, 4_8, and 8_8) had a positive effect on woody species survival (Figure 6).
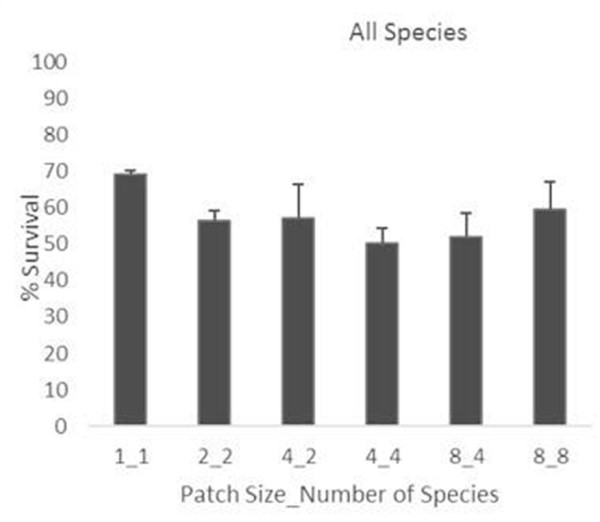
Figure 5 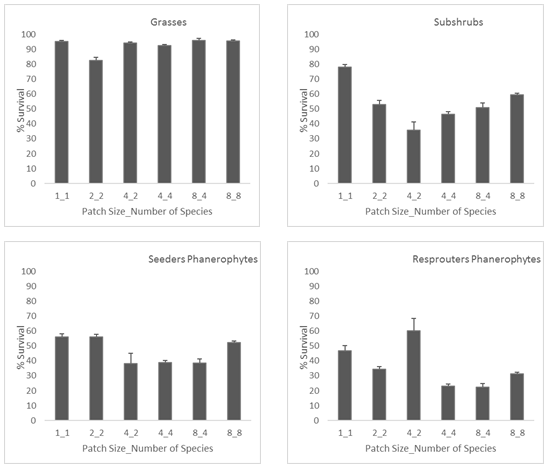
Figure 6 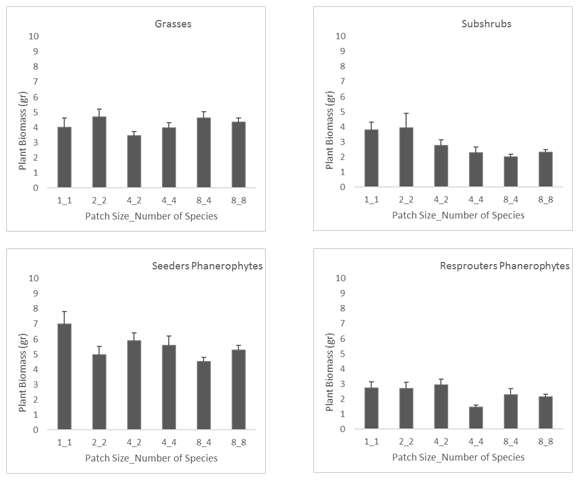
Figure 7
These results question traditional restoration approaches that assume that early colonists may inhibit the incorporation of new individuals, and promote the introduction of shrub and tree seedlings after the removal of neighbouring vegetation (Savill et al., 1997). Conversely, the results suggest a positive net outcome from the trade-off between a relatively low risk of decreasing survival and the unquestionable benefit of introducing high diversity patches in degraded drylands, which is expected to increase ecosystem functioning (Maestre et al. 2009), without significantly increasing restoration cost (James et al., 2013).
The variation in sapling biomass with increasing patch size and diversity was quite small (Figure 7), i.e., individual biomass was not significantly reduced by increasing the number of accompanying species in the same patch. This result further supports the idea discussed above about a positive and cost-effective net outcome resulting from introducing several species with contrasting plant features in the same plant patch in the framework of dryland restoration.
It must be stressed that the results presented here correspond to early stages in the restoration trajectory (one year after planting), and thereby some degree of uncertainty exists about further development of the restored vegetation. However, it is worth noting that the earliest stages in restoration are the most critical ones, as survival typically decreases early after plantation (or seed germination), but then stabilizes after very few years (Chirino et al. 2009). Also, we could expect a larger niche differentiation and reduced competition between species with plant aging, as rooting depth and other critical plant traits differentiate according to the species functional group, provided these are contrasting groups.
Conversely to our expectations, functional diversity did not appear to be more relevant than species diversity for plant patch performance at this early stage of the restoration trajectory. For example, differences between treatments 4_2 and 4_4, which imply increasing both species and functional diversity from 2 to 4, were of the same order than differences between 8_4 and 8_8 that only had an increase in number of species (from 4 to 8) but shared the same number (4) of functional groups. However, as discussed above, it may occur that the contrasting features that distinguish the species from different functional groups fully develop with plant aging.
Note: For full references to papers quoted in this article see