| Authors: | Bautista, S., Urghege, A.M., Morcillo, L., López-Poma, R., Camacho, A., Turrión, D., Fornieles F., Rodríguez, F. and Mayor, A.G |
| Editor: | Jane Brandt |
| Source document: | Bautista, S. et al. 2015. Report on the potential for sudden shifts. CASCADE Project Deliverable 4.1. 26 pp |
Critical questions and experimental design
The transfer of resources from bare-soil interpatches to downslope vegetation patches contribute to plant productivity and overall ecosystem productivity (Aguiar and Sala 1999, Yu et al. 2008, Turnbull et al. 2012). However, patch growth would contribute to reduce the size of the bare-soil areas, which in turn would reduce the amount of resources transferred to downslope patches. According to Puigdefábregas et al. (1999), in a functional ecosystem, the relative amount of source and sink areas remains within a certain range of variation around a hypothesized optimum source:sink ratio that maximizes both the availability of resources and the growth of vegetation patches. Using a spatially distributed model, Urgeghe et al. (2010) found that total amount of runon water reaching herbaceous patches was maximum at intermediate values amount of bare-soil cover, which points to a tradeoff between the source area for generating runoff and the sink area for capturing runon. Despite the increasing interest in the interactions between vegetation spatial pattern, productivity, and hydrological processes, there is still very scarce experimental fieldwork testing the underlying assumptions and quantifying the feedbacks involved (Asbjornsen et al. 2011). Furthermore, the scarce experimental and modelling works conducted on this topic hardly consider multiple species at once, ignoring the potential role of plant diversity as modulator of the ecohydrological feedbacks operating in drylands. In the framework of CASCADE, we designed a manipulative experiment that focused on the global feedback between plant pattern (and associated bare-soil connectivity) and resource conservation. The experiment specifically assessed (1) the role of plant pattern (patch density and size) on resource conservation for different plant species (2) the role of plant diversity in modulating pattern effect on resource conservation, and (3) the impact of resource conservation/loss on plant performance.
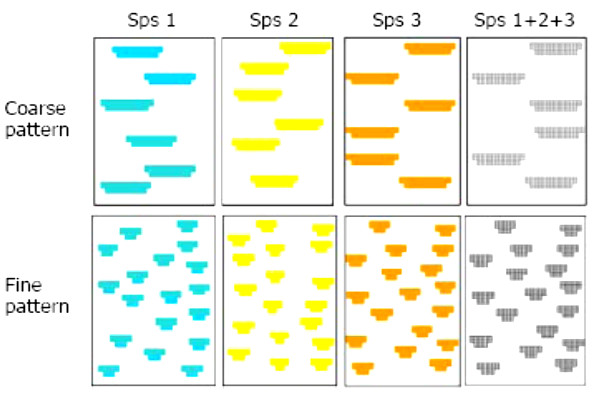
Using 1-year-old plant seedlings of three different species, we created artificial plant communities with contrasting plant pattern: coarse-grained pattern (6 large patches of 9 individual plants each), vs. fine-grained pattern (18 small patches of 3 plants each), and contrasting diversity: mono-specific vs. diverse communities. Total number of plants per plot was the same (54 plants) for all plots, and initial cover values were very similar. The species used were one steppe grass (Lygeum spartum L.) and two shrubs (Atriplex halimus L., and Phillyrea angustifolia L.). In the “diverse” communities, all the plant patches include the three species used (3 plants/species in the patches of the coarse-pattern plots, 1 plant/species in the patches of the fine-pattern plots). In the “monospecific” plots, all patches include the same single species. The plant communities were established on 24 plots of 1x2-m, located on an artificially created slope. There were 3 plot replicates for each treatment combination (2 spatial patterns X 4 plant communities). Over a one-year period, we monitored (1) Runoff and sediment yields from each plot, (2) Soil moisture in plant patches and bare-soil interpatches, (3) bare-soil connectivity, and (4) plant performance (survival, growth).

Hydrological response to plant pattern and diversity
Overall, the two contrasting plant spatial patterns assessed in this experiment (coarse and fine) did not show significant differences in the hydrological response of the experimental plots. This result is due to the fact that the differences in runoff and sediment yield between coarse pattern (i.e., few big patches) and fine pattern (i.e., many small patches) were not consistent across species, with Atriplex halimus and Lygeum spartum being more productive in coarse patterns and Phillyrea angustifolia being more productive in fine patterns.
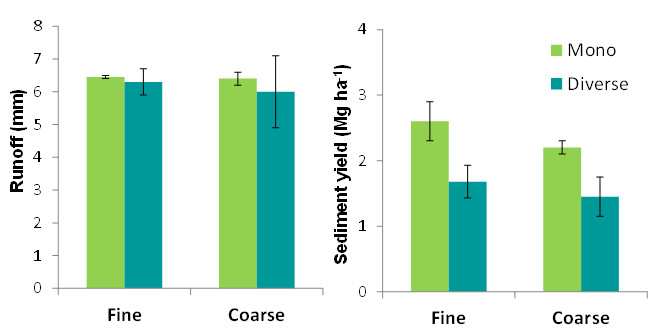
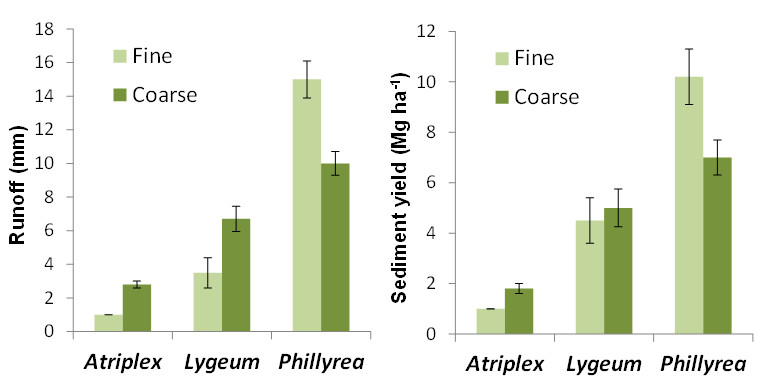
Plant diversity did not show any significant effect on runoff production, but it significantly reduced sediment yield. Thus, diverse plots produced less than average than monospecific plots, similar or even less than the least productive monospecific assembly (Atriplex), which suggest some kind of complementary of plant traits regarding the control of soil loss and the capacity for trapping and storing sediments. For example, while the dense canopy of A. halimus plants may have protected the soil from detachment by raindrop impact, the dense ground cover of L. spartum may have been particularly effective in trapping sediments that were transported by overland flow.
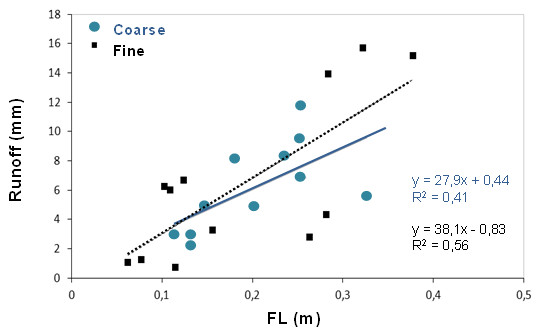
Regardless the broad class of plant spatial pattern of each plot, total plot runoff (Fig. 10) and sediment yield (data not shown) were positively correlated to the connectivity of the bare soil areas (measured with the Flowlength index; Mayor et al. 2008), pointing to this property as a key factor driving runoff and sediment yield in drylands. Although our initial expectations were that bare-soil connectivity would be greater for coarse-grained patterns than for fine patterns, both patterns resulted in a similar and rather wide gradient of bare-soil connectivity. This fact revealed that only when coarser patterns imply more connected bare soil, a negative impact of this type of pattern on water and soil conservation could be expected (e.g., Bautista et al. 2007).
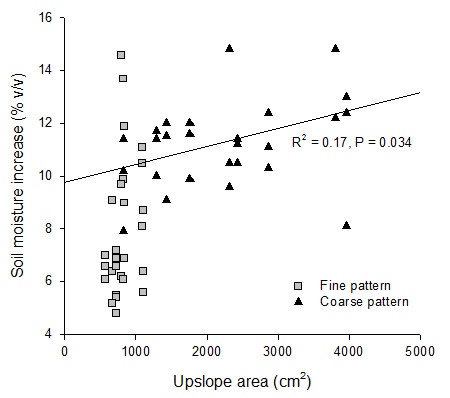
Finally, plot-averaged soil moisture was generally similar between coarse and fine patterns. However, at the patch scale, there was a consistent trend of higher water gains in the plant patches of the coarse pattern than in the patches of the fine pattern. One of the reasons to explain this behavior is higher runoff redistribution from inter-patches to patches in the coarse pattern due to larger runoff-contributing areas (i.e., bare soils) upslope the patches. This possibility is supported by the positive relationship between soil moisture increase after a large rainfall and the size of the upslope contributing area of the patches, only found in the coarse-pattern plots.
Overall, our results show that the key pattern property driving runoff and sediment yield is the connectivity of the bare soil, regardless the grain size of the pattern: coarse (i.e. few big patches) or fine (many small patches). However, big patches, with big bare-soil inter-patch areas upslope receive more runoff water after major rainfall events. These results highlight the interplay of local (patch-scale) and global (plot scale) effects of bare soil connectivity, and can be used to feed and validate dryland models that consider these effects (e.g., Mayor et al. 2013).
Vegetation response to plant pattern
The results reported in previous sections consistently showed that both plant cover and pattern are critical for resource conservation in semiarid patchy landscapes, even within a relatively small range of variation in plant cover. A critical question that derives from these results is to what extent this pattern-driven variation in resource availability determines plant performance and contributes to further variations in plant cover and pattern.
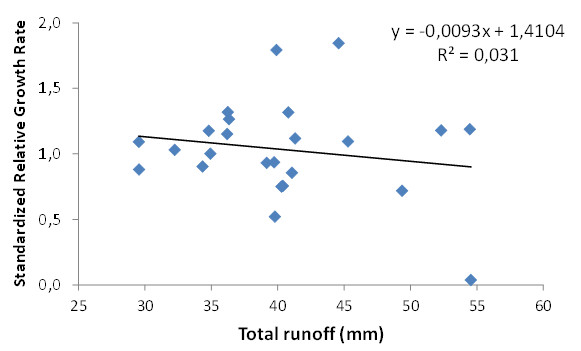
To address this question, we assessed how individual plant growth depended on the variation in runoff (and so in water losses) driven by differences in plant pattern and cover. We monitored 27 individuals from each plot and estimated the average standardized relative growth rate per plot by (a) calculating the relative growth in basal diameter, BD (Ln BD final – Ln BD initial) for each monitored individual in a given plot, (b) standardizing the individual growth rates by making them relative to the average growth rate of the respective species, and (c) averaging the standardized individual values per plot. The figure below shows the relationship between average standardized growth rates and water losses from runoff for each experimental plot. Although the data seem to follow a decreasing trend, no significant relationship was found between these two variables. Several factors may have contributed to this result. On the one hand, the spatial (2-m² plots) and temporal (1 year) dimensions of the manipulative experiment reported here have resulted in only small differences in water losses between plots. Thus, the largest difference between plots (25 mm of total runoff) represents less than 10 % of the total rainfall input (335 mm) during the study period. These differences in resource availability appeared to be rather small to influence plant growth at the global (plot) scale. On the other hand, plant performance is expected to also respond to variations in water inputs at the patch scale, and thus while larger bare-soil connectivity leads to larger runoff losses, it also implies larger water inputs at the patch scale, which may have counterbalanced and masked the impact of small differences in global losses. Overall, these results point to the need of further investigating the interplay between global and patch scale feedbacks between plant pattern and resource availability, upscaling the spatial (see »Patch-scale effect of resource redistribution on plant performance) and temporal extent of the experimental settings. On-going additional large-scale experiments in CASCADE address this need and investigate the long-term impact of vegetation removal (and associated changes in plant pattern and bare-soil connectivity) in plant pattern dynamics.