Introduction
In the case of dry Mediterranean ecosystems, one key mechanism is the interplay between plant-plant interactions at small and large spatial scale. At small spatial scales, local facilitation occurs when neighbouring plants “help” each other, by means of shading, the improvement of local soil quality through the build-up of soil organic matter, and/or the conservation and acquisition of water and nutrients. At large spatial scales, the plants compete for the same limited resources which constrain the growth of the population. This effect is especially pronounced under relatively harsh environmental conditions, where the contrast between bare ground and the ameliorated conditions in the direct vicinity of a plant is strongest, and were the overall scarcity of resources drives global competition. Facilitation is exerted through multiple mechanisms that, in general, imply improved soil conditions underneath and near plant patches, particularly higher water infiltration capacity (Mayor et al., 2009) and nutrient cycling (Mayor et al., 2016).
The interplay between local facilitation and global competition results in vegetation forming patterns, as plants in a patch with other plants will be better able to survive harsh conditions than alone (facilitation) while, at the same time, patches compete with each other (competition). Patterns that result from facilitation and competition can take several forms, for example gaps of bare soil surrounded by vegetation and labyrinths of vegetation and bare soil (Rietkerk et al., 2004).
With decreasing overall vegetation cover, the occurrence of a catastrophic shift becomes more likely (Rietkerk et al., 2004). Furthermore, while passing the critical threshold means that, even together, plants are not able anymore to survive, recovery through the establishment of single plants will require higher levels of resource availability, precisely due to the absence of facilitation, at least during the initial stage of re- establishment. For this reason, the avoidance of catastrophic shifts is to be preferred over rehabilitation as recovery can be ecologically difficult and economically expensive.
To understand ecosystem behaviour under stress, not only interaction between plants and plant species, but also interaction between plants and their direct environment, including soils, are important. A number of research efforts have tried to predict ecosystem shifts, using plant and soil indicators. The focus of the indicators has varied depending on the site study and main variable.
Kéfi et al. (2007) analysed spatial vegetation patterns in terms of the frequency distribution of vegetation patch sizes in three arid Mediterranean ecosystems in Spain, Greece and Morocco. They showed that the patch-size distributions away from tipping points follow a power law, while those approaching a tipping point deviate from a power-law distribution. These deviations correspond to a disproportionate high frequency of relatively small vegetation patches. The mechanism behind this is that the patches shrink to the minimum size at which they can survive through facilitation. In this way, spatial vegetation patterning can serve as an early-warning signal for the proximity of tipping points and unwanted, sudden catastrophic shifts.
CASCADE's results
Grazing stress-level experiment At four CASCADE sites, we assessed changes in soil quality and plant performance as a function of a grazing-stress gradient, with the aim to identify possible functional thresholds controlling critical shifts in the soil-plant systems.
In the Castelsaraceno and Santomera sites, soil quality parameters did not vary significantly between grazing-stress levels. These two sites represented the wettest and the driest CASCADE sites, respectively, and were subjected to grazing pressures that were, overall, low to moderate. These relatively low grazing intensities were the most plausible cause for the lack of effect of the stress gradient in both sites. A possible explanation for this lack of grazing impacts in these two sites might involve an interactive effect of climatic conditions, with the role of grazing being reduced under the most extreme, driest as well as wettest experimental conditions. The relative wet climate of Castelsaraceno, together with its high vegetation cover, might make its soil-plant system robust enough to deal with the current grazing levels. In the case of Santomera with its very dry conditions and reduced overall vegetation cover, one might expect that soil conditions are mainly affected by water availability, being the limiting factor for vegetation cover and growth and, ultimately, soil quality. Conversely, the remaining two sites, Messara and Randi, showed noticeable effects of grazing-stress levels on several of the measured soil quality indicators. At the same time, however, no clear pattern emerged as to how grazing-stress levels were linked to threshold values of soil quality indicators. However, overgrazing generally reduces the vegetation cover, which increases soil erosion and leads to a reduction in soil fertility and thus productivity. CASCADE studies have shown that if surface cover falls below 30-40%, soil erosion increases sharply because of the high connectivity of bare patches (Mayor et al 2016).
The comparison of the soil quality indicators of vegetation patches and inter-patches provided more consistent information. Topsoil organic matter and nutrient contents were clearly higher in vegetation patches than in inter-patches in two of the three study sites where the vegetation revealed a well- defined patchy pattern, i.e. in Santomera and Randi as opposed to in Messara.
The effects of the different grazing-stress levels on the performance of the targeted plant species differed markedly between the four study sites. In Santomera, canopy cover and basal twig diameter of Anthyllis cytisoides did not vary noticeably with grazing-stress level, although there was a tendency for twig growth to be higher at the high-stress plots, probably due to compensatory growth. In Randi, canopy cover and branch basal diameter of Calicotome villosa showed higher values for the low and intermediate stress levels than for the high stress level. In Castelsaraceno and Messara, neither plant cover nor biomass of the two target species Brachypodium rupestre and Hyparrhenia hirta, respectively, showed clear differences among the three grazing-stress levels.
A possible explanation for these findings follows a similar kind of reasoning as presented earlier, on the observed effects on soil quality. Grazing impacts are reduced not only in marginal ecosystems that are essentially controlled by lack of water availability (Santomera) but also in productive ecosystems without lack of water availability and high vegetation cover (Castelsaraceno), except perhaps if grazing pressures become much higher than at present. From the two sites with less extreme conditions Messara showed a noticeable impact of grazing-stress level on soil quality but not on plant performance, while Randi revealed a consistent grazing effect on both soil quality and plant performance.
Drought stress experiment No consistent effects of the enhanced drought periods on the measured indicators of soil quality and plant performance were found. For the driest sites, Randi and Santomera, we found increased nutrient availability and, to a lesser extent, higher carbon contents with increased drought stress. This is probably due to an increased amount of litter and dead roots being degraded (López-Poma and Bautista, 2014). Similarly, for Randi and Santomera we found a decreasing trend in plant performance with increasing drought stress, but not for the other two sites. These results indicate an extraordinary capacity of the plant-soil systems of these very dry areas to cope with drought, as only the combined effect of a severe natural drought plus the additional experimentally-induced drought finally produced a decrease in plant performance.
Plant pattern experiments Our findings (Bautista et al 2015) have demonstrated that both plant cover and plant pattern exert a critical role in controlling water and soil conservation in patchy ecosystems. This role mainly relies on the sink capacity of the soils underneath the plant patches, rather than on the capacity of the patches for rainfall interception and physical obstruction to overland flow, but both these phenomenon enhance water storage below a plant patch. The connectivity of bare-soil emerged as the most critical pattern attribute for explaining the hydrological behaviour of patchy ecosystems, as it reflects and depends on both cover and pattern. Larger bare-soil connectivity implies larger water and sediment losses from semiarid slopes, but it also implies larger inter-patch areas, which is beneficial for the performance of the downslope patch. Although plant cover and biomass are the most common vegetation properties used for hydrological modelling, our results suggest that other patch metrics like patch number and/or size distribution could be better hydrological indicators than patch cover. Integrated indexes based on capturing the connectivity of the bare-soil matrix in patchy ecosystems, such as Flow length index, have great potential as surrogates for the hydrologic functioning in semiarid landscapes. These indices can be easily obtained from aerial photographs and incorporated into hydrologic and erosion models at the hillslope and catchment scales.
Bautista et al (2017) showed that, at the patch scale, the effect of patch diversity and size on plant performance depended on the plant functional types considered and the environmental conditions, yet some common pattern was found for a large variety of dryland species tested.
At early stages of the restoration trajectory (first 1-2 years after planting), with all plant seedlings sharing similar rooting space, there was no evidence of complementarity between species that may have resulted in higher productivity in multispecies patches as compared with monospecific patches. However, there was no evidence either of detrimental effects of interspecific competition, as compared with intraspecific competition in monospecific patches. Big diverse patches benefited better from the higher capacity for trapping water and other resources from runoff than big monospecific patches. Under stressful conditions, facing both intra-specific and interspecific competition within the plant patch is more challenging for the species than interacting only with conspecific individuals.
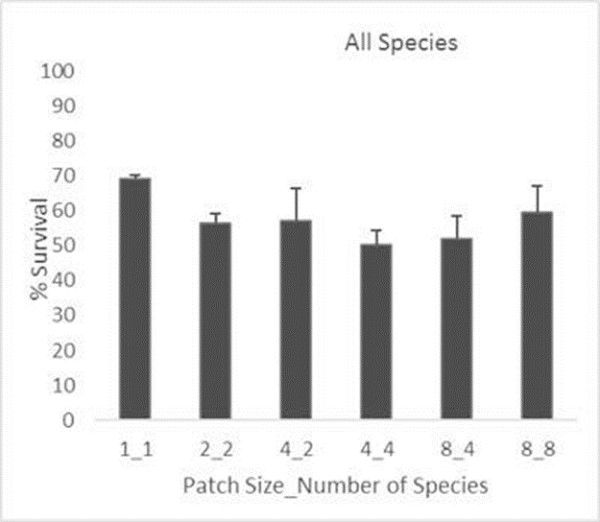
Figure 5
Compared with patches with a single plant, individual biomass was not significantly reduced by increasing the number of accompanying species in the same patch (Figure 5). Increasing patch size and diversity may reduce to some extent the probability of sapling survival in the restored patch. However, in general, the reduction in survival with increasing diversity is minor suggesting a positive net outcome from the trade-off between a relatively low risk of decreasing survival and the benefits derived from increasing diversity. Functional diversity did not appear to be more relevant than species diversity for plant patch performance at early stages of the restoration trajectory.
At the community scale, low initial plant cover did not constrain the potential for restoration success, which could be explained by the positive effect of water and sediment transfer from large bare soil areas to few existent plant patches. Our findings have demonstrated that eco-hydrological feedbacks between resource redistribution and vegetation dynamics that are mediated by bare-soil connectivity exert an important role in modulating the restoration potential of dryland ecosystems. Larger bare- soil connectivity implies larger water and sediment losses from semiarid slopes, but it also implies larger inter-patch areas and associated larger run-on inputs to existent plant patches, which is beneficial for the performance of the vegetation in the patch. This local feedback, if strong enough, increases the range of conditions (external stress, minimum initial cover) that allow the recovery of the system.
From an applied perspective, in a context of dryland restoration, a number of recommendations can be derived from our results, including
- using (creating) multispecies big patches, yet minimizing intraspecific competition by reducing the number of individuals per species within the same patch;
- spatially arranging plant patches on slopes in a way that maximizes the capture of runoff water by plant patches;
- combining species in the plant patches with plant traits that maximize the capture and deep infiltration of runoff water.
Modelling All the models developed in CASCADE confirm the importance of positive feedbacks in driving the emergence of catastrophic responses at the ecosystem scale as a response to increasing stress. We showed that incorporating a realistic aspect of grazing, which is that grazers tend to eat more at the borders of vegetation patches in a patchy landscape, decreased dryland resilience by increasing the size and the probability of catastrophic shifts under increasing stress (drought or grazing intensity) (Schneider & Kéfi 2016). We also found that incorporating a feedback between fire occurrence and vegetation composition promoted, in combination with drought, the emergence of alternative stable states and therefore of possible catastrophic transitions between those states (Baudena et al. In preparation). The connectivity-mediated feedback (i.e. the feedback between vegetation pattern, resource redistribution and productivity) decreased the amount of pressure required to cause a critical shift to a degraded state (Mayor et al. 2013). Not including these feedbacks into dryland ecological models may lead to an overestimation of ecosystem resilience and therefore failures in the prediction of catastrophic shifts.
Furthermore, the model results suggest that the upcoming climate change predicted for Mediterranean drylands, and in particular the rainfall patterns, could induce and enhance the occurrence of catastrophic shifts in those ecosystems.
The fire models suggest that the oak forests are very resilient and that in this case, catastrophic shifts may actually be less common than previously thought (Baudena et al. In preparation; Vasques et al. In preparation) (although alternative states emerge when a positive feedback between vegetation composition and fire occurrence is introduced in the model ; Baudena et al. In preparation).
The models developed contribute to the fundamental understanding of what determines the species composition of a given dryland and how that drivers the response of the ecosystem to increasing stress, in particular: why and how alternative stable states, and therefore catastrophic shifts, occur in drylands. This fundamental understanding provides some keys for 1) preventing dryland degradation and 2) restoring degraded ecosystems. The grazing model (Schneider & Kéfi 2016) suggests that vegetation patterns provide early warning signals of approaching desertification (i.e. the spatial structure itself). However, spatially heterogeneous grazing does not only altered ecosystem stability (by increasing the probability of catastrophic shift) but could also blur the early warning signals at high grazing pressure. This suggests that we need to be cautious regarding the use of early warning signals of ecosystem degradation when the pressure at play has a spatially-explicit component. It also suggests that additional indicators of degradation need to be developed taking into account the spatial component of the stressor.
Furthermore, model simulations suggest that using a bare-soil connectivity index (Flow length; Mayor et al. 2013), in addition to vegetation cover and pattern, may provide more informative early-warning indicators of dryland degradation. Mayor et al. (2013) concluded that bare-soil connectivity and vegetation patterns both form important early-warning indicators for dryland degradation, Verwijmeren et al. (2014) found that the aspect of hillslopes has the largest influence on vegetation cover and soil functioning, furthermore they found species association strength to be an important factor at both species pair level and community level for maintaining ecosystem functioning. Tirabassi et al. (2014) found that certain vegetation pattern metrics may offer possibilities in identifying an upcoming critical transition in semi-arid ecosystems. They also introduced new measures to assess the quality of these indicators, and found new early warning measures to be of higher quality than classical indicators. Mayor et al. (2016), researched the quantity and quality of soil organic matter in forest areas with different fire frequency regimes, and found that a year after a fire, the amount of soil organic matter increased, while its quality was lower (expressed as the ratio of labile and total organic) and areas undergoing repeated fires decreased their likelihood of recovery due to the high loss in resources. The researchers concluded that the labile organic matter fractions in the soil might be used as an early-warning indicator for shifts in soil fertility in response to fire recurrence.
Eventually, quantifying those indicators derived from model studies on field data may help identify field sites that are at the higher risk of irreversible degradation and prioritize those for conservation measures.
For a given ecosystem studied, the models developed allow reaching a better understanding of how different interactions and drivers control the composition of dryland communities and their changes through time. Such a knowledge can be extremely useful in terms of management, e.g. to foster one given community over another (or to prevent being trapped in an undesired community). For example, the model results underline the importance of the practice of planting seedlings from late successional, resprouting species to increase the resistance and resilience to forest fires (Valdecantos et al 2016). The modelling approach reinforces such practices as it underlines the importance and the resilience of late successional resprouter species on the time scale of a few generations of these plants, which is well beyond human observation.
Moreover, the essential role of facilitation for both species coexistence and ecosystem resilience highlighted by the models suggests that it may provide a good opportunity for ecosystem restoration. In a degraded dryland, remaining adults individuals can be used as nurses to increase the recruitment probability of seedlings planted below or close to their canopies. In degraded grazed drylands, the same strategy can be applied using preferentially nurse species adapted against grazing, to improve the early survival of the planted seedlings.
Discussion and conclusions
Most of the empirical results obtained by CASCADE showed that increased stress through grazing (stress-gradient experiment) or drought (drought-experiment) deteriorated the plant-soil ecosystem, probably moving it to critical points for catastrophic shifts to happen. Since we did not actually observe such a catastrophic shift taking place in any manipulative rainfall-exclusion experiments, we cannot say where these tipping points lie. However, in the case of the Randi site on Cyprus, the increasing contrast in soil conditions between patches and inter-patches with increasing level of grazing pressure pointed to the proximity of a critical shift into a degraded state. Results also indicated positive feed- backs and local facilitation, as most of the statistically significant differences in soil quality and plant performance corresponded to better values for vegetation patches than for inter-patches. These results suggests that shifts would involve changes in both vegetation and soil, and that indicators reflecting both vegetation pattern and soil quality could provide early warning signals of shifts. Understanding of plant-soil and plant-plant interactions is also crucial to understand shifts in Mediterranean dryland ecosystems. Such understanding can improve predictions of shifts, but can also be used to increase the chance of success of restoration efforts.