| Authors: | Alejandro Valdecantos and Ramón Vallejo (CEAM) with input from study sites |
| Editor: | Jane Brandt |
| Source document: | Valdecantos & Vallejo. (2015) Report on structural and functional changes associated to regime shifts in Mediterranean dryland ecosystems. CASCADE Project Deliverable 5.1. |
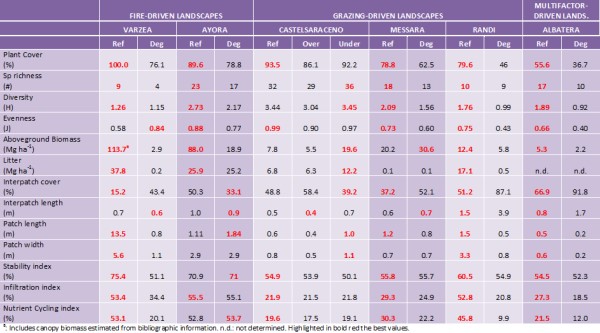
CASCADE field sites include different Mediterranean ecosystems in a wide range of climates and degradation pressures and, as a consequence, the range of values of ecosystem properties obtained is also very large. Although comparisons between such different sites is not straightforward, Albatera, which holds the lowest aridity index, and, in a lesser extent, Randi represent the ecosystems with lower quality and functioning. Albatera presents the worst values of plant and interpatch cover, patch size (both Reference and Degraded states), diversity and infiltration (Degraded) and stability indices (Reference) of all six sites. On the contrary, Ayora offered the highest values of the Degraded states of all sites in plant spatial distribution and LFA indices. Biodiversity indices were highest in Castelsaraceno both for the Reference and the Degraded (Undergrazed) states and lowest in Várzea (Reference) and Albatera (Degraded). Várzea showed the two extreme values for aboveground biomass: the lowest in the Degraded (just two years after the last fire) and the highest in the Reference. In general, climate conditions frame degradation vulnerability through their effect on plant productivity and, probably, in plant community resilience to disturbances.
Areas under pressure from grazing
In general, the plant communities in the Degraded situations are very different from the respective healthy References both in composition and abundance. Pressure resulted in more homogeneous communities than in undisturbed states, except in Randi, where high heterogeneity is observed within both Reference and Degraded ecosystems, and Castelsaraceno, with little variations within degradation levels. Intense grazing may represent a stronger effect than soil texture in determining vegetation pattern distribution (Fuhlendorf and Smeins 1998). However, Adler et al. (2001) suggested that grazing might lead either to higher or lower heterogeneity depending on both the pre-grazing spatial pattern of vegetation and grazing.
Our field sites affected by grazing showed a generalized decrease in diversity with grazing pressure and, hence, can be described as overgrazed (Perevolotski and Seligman 1998). Papanastasis et al. (2015) applied the same methodology to assess the change in ecosystem services due to grazing in rangelands in Greece. They obtained very similar results to those we observed in CASCADE grazing sites, with a significant loss of biodiversity, stability, infiltration and nutrient cycling indices, plant cover and size and cover of vegetated patches in heavily grazed areas. But they also reported increases in some properties and ecosystem services when grazing pressure is moderate. Some authors have observed an increase in plant species diversity when disturbed by grazing (Belsky 1992), partly because of the change of competitive relationships between plant species (Crawley 1983). In addition to these change in diversity, we observed a profound change in species composition in all grazed sites, more modest in Castelsaraceno, as demonstrated in the PCA analysis and figures. Holocheck et al. (1989) reported a shift in species composition due to heavy grazing, partly due to an increase in the competitive ability of unpalatable species. But many dominant species of grazed rangelands present morphological and biochemical mechanisms to withstand grazing providing a relatively high resilience to the system (Perevolotsky 1995). Aboveground biomass was reduced in two out of three grazed sites (Castelsaraceno overgrazed and Randi) but increased in Messara. The reduction in leaf area associated to grazing severely affects to primary production in herbaceous pastures such as Castelsaraceno but the same does not happen in areas where shrubs are more abundant such as Randi. Shrubs have longer annual growth cycles and allocate photosynthates to woody parts and roots, often showing compensation growth responses (Tsiouvaras et al. 1986; Perevolotski and Seligman 1998). However, in sites that present a long history of grazing, higher levels of pressure may not significantly affect productivity. Plant pattern in the grazed states is markedly different than in the ungrazed ones, with higher interpatch cover and lower length and width of the plant patches in the grazed plots. These changes reduce the resource sink capacity as observed in other areas subjected to grazing (Papanastasis et al. 2015). Similarly, LFA derived indices are lower in all Degraded sites than in their respective References suggesting a reduction of soil surface conditions and, hence, soil, water and nutrient conservation in the system (Papanastasis et al. 2015). Ecosystem services have shown important losses due to grazing in the order Randi>Messara>Castelsaraceno following a decreasing order of aridity. Wang et al. (2014) established 0.32 as the threshold value of the aridity index that determines net N losses or accumulations. Castelsaraceno and Randi are well above and below this value of aridity, respectively, while Messara is just in that supposed threshold. On the other hand, grazing, especially when intense, represents an important tool to reduce fire risk in areas like the Mediterranean basin with prolonged drought periods by reducing the amount of fuel susceptible to burn during them. In these cases, grazing systems provides another service to people as it is the reduction of fire hazard.
Biodiversity and ecosystem functions in drylands have been observed to depend on the relative cover of woody species, with linear relationships in dry-subhumid sites and hump-shaped curves peaking at relative woody covers of 40-60% in semiarid (Soliveres et al. 2014). Isbell et al. (2015) suggested that the reduction in biodiversity generates an 'ecosystem service debt’ and defined it as 'a gradual loss of biodiversity-dependent benefits that people obtain from remaining fragments of natural ecosystems'. These authors highlighted the relevance not only of the extension of ecosystems but also the necessity of preserving and enhancing their quality in order to guarantee a sustainable provision of ecosystem services.
Areas under pressure from fire
The sites with fire pressure offer two complementary pictures of secondary succession after wildfires: a very initial stage of vegetation recovery in Várzea and a mature continuous shrubland without tree-canopy recovery in Ayora. In the short term, the ecosystem shows important reduction in species richness, biomass, vegetation patches, stability, infiltration and nutrient cycling. All these things result in an overall significant loss of ecosystem services. But this maritime pine forest system has the ability to recover with time most of them, if not all. Thirty-five years after the fire, the Ayora burned areas recovered ecosystem functionality to values of the Reference pine forest, and showed a spatial arrangement of vegetation that better conserves the resources, and accumulated similar amounts of understory and belowground biomass and litter. Pine regeneration after fire depends on many factors such as fire-interval, pre-fire basal area, slope aspect, land use history or competition with grasses at the seedling stage (Pausas et al. 2003, 2004; Baeza et al. 2007). The scarce presence of pines in the Degraded states of Ayora field site resulted in a significant reduction of the C sequestration service and could be improved by appropriate post-fire management.
The observed shift from forest to non-forest (shrubland) vegetation observed in Ayora is not uncommon especially in drylands. The very high fire recurrence of the Degraded plots in Várzea and the short interval between the two latest fires (2005 and 2012) may cause the change from forest to non-forest vegetation in this area as the time for the first flowering in Pinus pinaster may take between 4 and 10 years (Tapias et al. 2004). This imbalance between fire regime and dominant plant species’ life histories or unfavorable post-fire conditions may result in a failure to recover pre-fire carbon stocks and hence C sequestration service (Rocca et al. 2014). Stephens et al. (2013) suggested that this shift might not be catastrophic but would affect most ecosystem services. All ecosystem services showed significant short-term losses after the fire (Várzea) but only biodiversity and C sequestration losses lasted in the long term (Ayora). However, the particular conditions in Várzea, especially the higher water availability (0.84 and 0.26 aridity indices in Várzea and Ayora, respectively) suggest that the recovery of these assessed ecosystem services will be faster.
Area under multiple, diffuse pressures
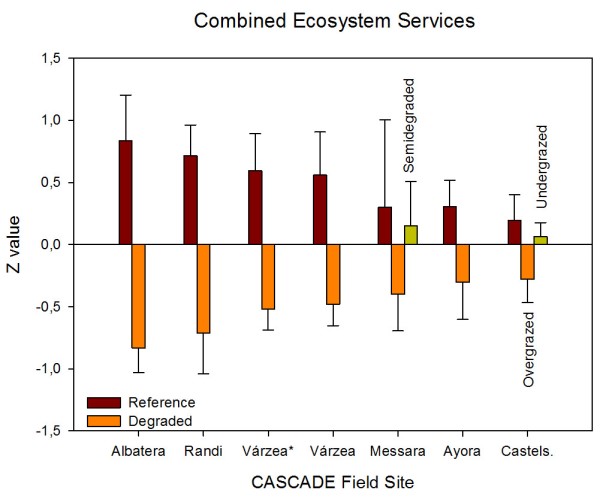
Albatera showed the highest relative losses of all individual ecosystem services and the combined value of all CASCADE field sites. It is the most stressed site as reflected by the very low aridity index (0.16 classified as semi-arid) and multiple diffuse pressures are and have been acting in the place for long. The main ecosystem properties affected by degradation were those related to the spatial distribution of vegetation and open areas (sink/source spatial pattern). The Degraded landscape showed a reduction of vegetation cover, with less and smaller patches of vegetation at longer distances from each other, and higher proportion of bare soil, which in turn reduces capacity of water infiltration and nutrient cycling, and decreases water conservation and soil conservation, and, finally, reduces productivity (Boeschoten 2013). The loss of water conservation is said to be the most essential function in semi-arid ecosystems (Whitford, 2002). Biodiversity was also highly reduced in the Degraded areas probably related to the absence of tall shrubs that act as keystone species in these semiarid shrublands (Maestre and Cortina 2004). Rey Benayas et al. (2009) observed a positive relationship between biodiversity and ecosystem services and suggested that restoration efforts should be directed to increase biodiversity. Stability is the index that showed the lowest loss in the Degraded as compared to the Reference state. Previous works in semiarid Mediterranean areas have showed that the Stability index is less sensitive than the other LFA indices to detect differences between land uses and/or degradation levels (Mayor and Bautista, 2012). However, it could also be that current erosion in the degraded state is actually low due to accumulated effects of past erosion. Thus, higher surface stone cover due to past soil loss may be protecting the soil from further severe erosion. Similarly, higher surface compaction due to accumulated disturbance and the loss of the top soil layers, richer in organic matter, may result in higher runoff but lower soil loss (Mayor et al. 2009). López et al. (2013) found lower values of the LFA stability index as degradation increased associated to lower vegetation cover and patch density, length and width, but a further increase of the index with more intense degradation as the exposed rock surface is higher and the sediments susceptible to be transported is lower. These results could therefore suggest that the system might have overcome a threshold of irreversibility (Scheffer and Carpenter 2003).
Summary
In summary, degradation pressures severely impacted ecosystem properties and services of the selected ecosystems along the Mediterranean Basin in a wide range of ecological, biogeographical and historical characteristics. The higher the aridity, the higher the loss of ecosystem services. Some observed changes from the Reference towards the Degraded states suggest that certain degradation thresholds might have been passed.
References
See the full report for details of all the other work cited on this page