| Contributing Authors: | Sonia Kéfi, Florian Schneider, Angeles G. Mayor, Alain Danet, Marina Rillo, Simon Benateau, Jacob Keizer, Ana Vasques, Susana Bautista, Paco Rodriguez, Alejandro Valdecantos, Jaime Baeza, Ramón Vallejo, Max Rietkerk, Mara Baudena, Mart Verwijmeren, Koen Siteur, Rubén Díaz-Sierra |
| Editor: | Jane Brandt |
| Source document: | S. Kéfi, A. Vasques, F. Schneider, M. Rietkerk, A.G. Mayor, M. Verwijmeren, R. Diaz-Sierra and M. Baudena. 2016. Response of Mediterranean drylands to increasing pressures. CASCADE Project Deliverable 6.1, 54 pp |
In the Mediterranean basin, there is a long history of exploitation and deforestation, which resulted in the conversion of the natural landscapes to agriculture [27]. In the past decades, Mediterranean forests have experienced intense changes, which resulted in extensive afforestation using pine plantations, or in marginal cropland abandonment [28–30]. In turn, these changes have led to an intensification of the fire regime, with an increase in the amount of burnt area and in the frequency of wildfires [28,29].
Most Mediterranean plant species are well adapted to fire, commonly responding to it through different mechanisms. Their three main post-disturbance regeneration strategies consist of:
- plant survival through the resistance of below-ground buds to fire, followed by resprouting of the above-ground tissues (obligate resprouters, e.g. oaks),
- plant mortality but survival of the seeds (obligate seeders, e.g. pines),
- a combination of both these mechanisms (facultative species; [30,31]).
Studies of post-fire dynamics have shown that resprouter species (which are usually late successional) are more resilient to both short and long fire frequency intervals than seeder species [32]. Pine seeds are often enclosed in serotinous cones, which open right after the fire, allowing pine population recovery [33]. However, as most pine species do not resprout, the resilience of pines to frequent fires is limited by seed production. Thus, the occurrence of repeated fires in a short interval of time or of a single fire in a young pine forest can cause the local extinction of pines [33,34]. This eradication gives place to the dominance of other species, which could either be early or late successional species. Intensive use of the land and high wildfire frequency can favor the replacement of pine and oak forests by fire-prone shrublands [34,35]. Furthermore, the re-introduction of late successional species, such as oaks, in fire-prone shrublands can be limited by several factors related to seed dispersal and micro-climate conditions [36,37]. These recruitment limitations could arrest successional processes, locking the system in a shrubland state [35,36]. However, there is also evidence that oak species can become dominant in former pine forests that have been eradicated as a result of recurrent fires [32,38].
Species from various successional stages can co-occur after disturbance, and the time after disturbance at which they attain dominance depends on their growth rate and longevity [39]. High fire frequency could thereby either result in the dominance of seeder species with short life spans and persistent seed bank, or in the dominance of resprouter species with high growth rate, whereas fewer fires would favor the dominance of seeder species with longer life spans, such as pine species [40]. Even so, the abundance of obligate seeder species with short life spans can be compromised by recurrent fires, through the exhaustion of their soil seed bank [41,42]. Such circumstances could be advantageous for species with resprouting ability (facultative species and resprouter species).
- For the Várzea site, we developed a cellular automaton model to study the effect of fire succession in pine stands, and we focused on the effect of fire severity and on the occurrence of safe germination microsites, connected to the existence of a litter layer.
- For the Ayora site, we developed a spatially-implicit model for vegetation cover including six different vegetation functions, and we focused on the effect of the feedback between species composition and fire frequency, and on the combination of fire and drought effects. This second model has been calibrated with data from CASCADE and from previous EU projects EUFireLab and LUCIFER).
1. Effect of repeated fire in pine plant communities, Várzea (Portugal)
The conditions under which a given species composition is expected in Mediterranean areas with recurrent fires remains largely unknown. In addition to the factors already mentioned above, plant succession is expected to be also modulated by factors such as fire severity [43], seed sources and disperser preferences [37], and the occurrence of safe germination microsites. In particular, the existence of a layer of litter after a fire modifies the probability of plant colonization, and the characteristics of the pine litter layer after a fire are largely determined by fire severity [44,45]. Despite the important role of litter as a modulator of the composition and structure of plant communities, its effects have not been included so far in the analysis of the successional dynamics of pine stands under different fire regimes.
- explicitly takes the litter layer into account, and
- simulates the population dynamics of three plant functional types: pines, oaks (late successional species) and obligate or facultative shrubs (early successional species) [46].
This model was used to investigate how the wildfire frequency conditions affect plant succession dynamics in pine forests.
Using this model, we studied the long-term vegetation dynamics of the ecosystem under different fire frequency conditions, and we analyzed the dynamics of the transient period (medium-term), by investigating how long it takes for the late successional species (oak) to attain relative dominance. Additionally, we studied how plant species composition, colonization capacity (represented by the arrival of acorns) and post-fire microsite conditions (represented by fire severity and litter depth) influence succession, measured by the time when oak attains relative dominance.
We hypothesized that, in the long term, pine forests would be replaced, through facilitation processes, by late successional species (Quercus spp.) in undisturbed regimes. Moreover, increased fire frequency in pine forests where facultative shrub species are abundant would enhance the dominance of this type of shrubs (because of their resprouting capacity) and delay succession towards mature forests.
Model description
The model is an interaction particle system, where each cell is dominated by one of three plant functional types: pine, oak or shrub [26,46]. Accumulation of litter is a spatial-explicit process, that takes place in the cells occupied by a mature pine tree as well as in their eight neighboring cells. The different functional groups have different colonization probabilities of empty sites, which depend on the number of seeds available in the cell for each of the functional groups, the establishment capacity of each functional group, and the microsite conditions, determined by litter depth. The accumulation of a litter layer in the absence of disturbance is expected to have a prominent role in successional processes [47]. The germination of small seeded species under thick litter layers may be prevented [48,49], whereas bigger seeds tend to remain over the soil surface and germinate easily in such conditions. This pattern is observed consistently in Mediterranean resprouter (late successional) and obligate seeder or facultative shrub species (early successional), with the former having often bigger seeds and, at the same time, exhibiting less persistence in the soil [50]. Moreover, in pine forests, litter acts as a function of canopy cover, which by shading and reducing climate extremes and increasing water availability in the summer, favours the establishment of late successional species over early successional species [51–53]. Therefore, the model assumes that a higher litter thickness favours the relative dominance of late successional species while, at the same time, inhibiting the colonization of early successional species. Fire disturbance regime was defined stochastically, with the occurrence of fires defined by an exponential distribution of return times [54]. The simulation was first run for 100 years without fire. After that, we introduced fire with one of three different average return times: seven, fifteen and thirty years. We also kept a scenario without fire. These fire regimes were indicative of what was observed in the centre-north of Portugal, where the average fire frequency is 24 years, and where the regeneration of pines is threatened in 15% of the area [55]. When a fire occurs, the functional groups with resprouting ability (oaks or facultative shrubs) continue to occupy the same cell, whereas pines and obligate seeder shrubs die. In the high fire severity scenario, all the needles and litter are consumed, whereas in the low severity scenario, the needles of mature pines remain intact and fell on the soil surface, creating a litter layer.
Results
In the absence of fire, we found that the ecosystem reached a stable co-existence between the three functional types after about 1000 years, and this was independent of the initial conditions of the simulation. This steady state consisted in the dominance of oaks (~75%) with co-existing pines (~15%) and shrubs (<5%) (Figure 1). The changes in plant species relative dominance over time were related to the life span of each species: during the first decades, the short lived species (i.e. shrubs) dominated (after ~100 years), then the intermediate lived species (i.e. pines) dominated, and finally the long lived species (i.e. oak) dominated (at ~250 years). The relative dominance of oaks at steady state (i.e. cover values higher than 50%) was maintained under all fire frequency conditions (Figure 1). The co-existence of all three functional types, however, was not observed under fire disturbance, since pines went extinct in all fire frequency conditions. In case of high fire frequency and severity, obligate seeder shrubs also went extinct. When facultative shrubs were included in the simulations, a long term co-existence of shrubs and oaks was reached under all the fire disturbance regimes (Figure 1).
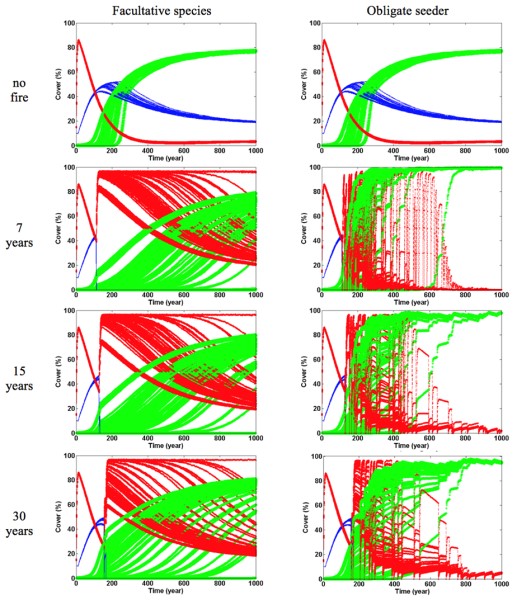
The time at which pines went extinct depended exclusively on the recurrence interval of fire events (Figure 1). However, the model results suggested that the time at which oak reached dominance depended mostly on the acorn yearly input, and thus on oak nearby populations. In particular, Oak dominance was delayed substantially (up to 800 years) when acorn influx rates were low. Moreover, high fire frequency accelerated oak dominance when coexisting with an obligate seeder shrub, but delayed it when coexisting with a facultative shrub species (i.e. with a resprouting ability). This result was observed for all influx rates of oak seeds and fire severity did not change this pattern considerably.
Conclusion
Our model results are in agreement with paleobotanical records, which suggest that the prevailing vegetation in the Iberian Peninsula during the past centuries or millennia probably consisted of a mixed pine-oak woodland, with a relative dominance of late successional species, such as Quercus robur [56]. The persistence of pines in the system has been linked to fire occurrence [57], which usually results in open areas that are favourable for pine colonization, but could also be explained by local less favourable environmental conditions for the regeneration of oak, such as steep slopes and cold or dry sites [58]. Furthermore, crop abandonment and the fact that very little artificial oak plantations have actually occurred limited oak seed sources, making acorn inputs close to inexistent in many instances, hence absolutely changing pine forest post-fire succession.
As expected, in the model, oak dominance under optimal conditions for oak establishment was accelerated by a higher yearly input of acorns. This could have important management implications since by understanding and manipulating the potential and effective dispersal of oak plants, we may be able to predict the time at which oaks would reach dominance. The main seed dispersals of oak species are jay (Garrulus glandarius) and mice, with the first having a much wider territory range (up to 100 ha) than the second (up to 1 ha; [59]). Furthermore, the species present in a given plant community can affect the colonization capacity of oaks, since, for example, pine stands foster seed arrival, whereas shrublands are usually avoided by jay, because of their higher exposure to predators in these open areas [60]. If seed arrival is inhibited by shrub dominance, the system can be locked in an early successional phase, especially when pines are eradicated from the system. This would agree well with the findings of Acacio et al. [36] who found seed arrival to be one of the limitations of oak establishment in shrublands.
2. Effect of recurrent fire and drought on vegetation succession in Mediterranean forest, Ayora (Spain)
We studied two more elements that are very important for Mediterranean forests. The first is the fact that some species are more prone to fires than others, as they have different e.g. flammability, heat and water content, standing dead biomass [61–64] (see »Structural and functional changes associated with regime shifts and »Restoration potential for preventing and reversing regime shifts). This determines the possible existence of a feedback between fire occurrence and species composition, as the most fire prone plant types are early successional shrub seeders, or grass resprouters, that can regrow very fast after a fire, and thus maintain the system in a highly fire-prone condition, while the slow-growing late successional species (mainly oaks) are much less prone to fires. As it has been shown in other ecosystems, e.g. in savannas, such feedback between fire and vegetation can create bistability in the system [65,66], and thus possibly determine the occurrence of catastrophic shifts in the vegetation composition.
The second element is drought and its interaction with fire, which is expected to increase the chances of a (possibly catastrophic) shift in species composition. For the Mediterranean area, an increase in temperature and in drought occurrence is expected [67]. Drought affects directly species composition, via the limitation of recruitment, growth and survival, to which the late-successional resprouter species are more subject than shrubs and pine seeders [68–73]. Drought also increases the probability of fire, thus also indirectly influencing species community composition. The combination of these direct and indirect effects can influence dramatically species composition and ecosystem functions [74,75].
Under the current climatic conditions, we expected, as in the study by Vasques et al. [46], that late successional species (Quercus spp.) would replace the other species, when given enough time. Moreover, we investigated the possible existence of alternative states between open shrubland and closed oak forest, due to the fire feedbacks, which are a necessary condition for catastrophic shifts to occur [77].
Model description
Inspired by the vegetation of the Ayora Cascade site (see also »Ayora, Spain: Structural and functional changes), the model represents six plant types, namely (in order from late to early successional, i=1...6 in Table 1): Quercus spp, Pinus halepensis, Rosmarinus officinalis, Ulex parviflorus, Cistus spp., and Brachypodium retusum. Oak (Quercus) and grass (Brachypodium) can resprout after fires, while the other four plant types (pines and three shrub seeders) regrow via seed germination after a fire.
Table 1: List of vegetation types in their hierarchical order
| i | Vegetation type (genus or species) | Acronym | Fire strategy |
| 1 | Quercus (ilex, coccifera) | Q | Resprouter |
| 2 | Pinus halepensis | P | Seeder |
| 3 | Rosmarinus officinalis | R | Seeder |
| 4 | Ulex parviflorus | U | Seeder |
| 5 | Cistus (mostly albidus, but also some monspeliensis) | C | Seeder |
| 6 | Brachypodium retusum | B | Resprouter |
The model represents the dynamics of the vegetation cover in a plot. We modified a model that was first introduced by Tilman [78] to study succession, in which we included specific terms to represent the responses of the different plant types to fire. The dynamical variables, bi, represent the fraction of space (0≤bi<1) in the plot occupied by a certain vegetation type i. After a fire, resprouters keep most of their cover (to represent the fact that they resprout out of their roots). Seeders, however, burn down completely, meaning that the space they occupy is reduced to zero, but their germination ability in bare soil is high after a fire, because of the seed bank originated before the fire. For pines (i=2), the seed bank falls from the canopies to the ground at each fire, and the seeds are then available for approximately two years. The seed bank depends on the age of the pines before the last fire, because pines produce seeds only at mature age (around 10-12 years). For shrub seeders (i=3…5), the seed bank is re-calculated at each fire as a function of the seed bank produced since the last fire (which is a product of the average plant cover between the fires and the seed production), the seed durability in the soil and the seed survival rate to fire.
Fires are modeled as stochastic events with an average return time [54]. We perform two types of simulations. In the first ones, there is no dependence of fire return time on vegetation cover. These runs are thus called the “no feedback” runs (and are similar to Vasques et al. [46] model runs). The second type of runs includes the feedback between fire and vegetation composition (the “feedback” runs). We assume that fire return time depends on vegetation cover and composition (similarly to e.g. [79,80]) such that a larger extent of more flammable species increases the average fire frequency.
The datasets
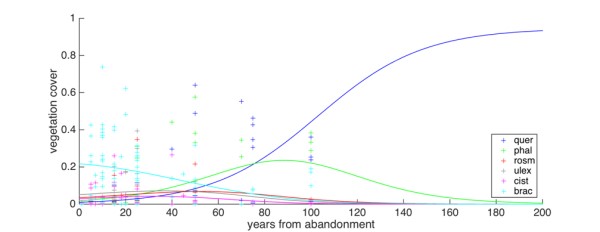
Succession: The model without fire was calibrated using data from different sites where vegetation cover had been previously recorded. Data were obtained in abandoned old fields ranging from 1 to 100 years since abandonment. A total of 75 sampling sites were used from previous studies on vegetation succession located in the Western Mediterranean Basin [35,81–84]. Data were a product in parts of CASCADE (Ayora site) research, and in other parts of the EU-project EUFireLab (FP4 EU project EVR1- 2001-00054 “Euro-Mediterranean wildland Fire Laboratory, a "wall-less" laboratory for wildland fire science and technologies in the Euro-Mediterranean region”, 2002-2006).
Fires: The data for calibrating the model including fires were obtained from a series of experimental sites located in SE Spain [85]. Data originated in parts from CASCADE (Ayora site), and in other parts from the EU-project LUCIFER (FP4 EU project DGXII ENV4-CT96-0320 “Land use change interactions with fire in Mediterranean Landscape”, 1996-2000). We used data from four sites, namely Onil, Pardines, Ayora (CASCADE), and La Torre. These sites have experienced several wildfires during the last three decades, and within the same site we dispose of three neighboring plots with 1, 2 or 3 fires. These plots are part of a permanent long-term study established in 1994 and they keep being sampled approximately every six months the first two years after a fire, once a year until 5 years after fire, and afterwards every 5 years.
Model calibration
We performed a model calibration to find the optimal parameters by minimizing the residual sum of square of the model trajectories and data. More precisely, we minimized the mean square deviation between model and data, to the variance of the data, using the index H2, that is zero for a theoretically perfect model, and is one when the variance of the data and the mean square deviation are equal [35,73–76]. The optimization was performed using the simulated annealing algorithm [87]. First, we calibrated the model using the successional data and without including fire in the simulations. We obtained an optimal value for the six colonization rates, and we also optimized the initial conditions for the six vegetation types (see model trajectories in Figure 2). Second, we calibrated the full model using the fire data. For each site, we used the historical fire sequence to determine time of fires and time between fires. As initial conditions, we used the first data in the time series (i.e. the vegetation cover at the time the second fire occurred, for all cases but Ayora).
Results
Although coexistence is theoretically possible in the model adopted for succession [78], with the set of parameters obtained from the optimization procedures, there was no possibility of coexistence in the long term, in the model without fire: oaks became dominant (here defined as their cover is above 50%) after about 120 years from abandonment, and all the other species disappeared within the first 150-200 years (Figure 2).
When fires were included in the simulations, vegetation covers did not reach steady state, but kept varying as a consequence of the stochastic fires, with the resprouters decreasing only of a certain fraction and the seeders becoming nearly extinct at each fire. Despite the fire disturbances, oaks eventually dominated the system, independent of the initial conditions or of the fire frequency, given that they have enough time to establish (Figure 3, 4 first column). The time at which oaks became relatively dominant depended, as expected, on the initial conditions of the oak itself (as in the model of Vasques et al. [46]), but not on the initial conditions of the other vegetation types. More frequent fires could slightly delay the oak dominance (Figure 3). The species composition in the initial part of the simulations, before the oaks became relatively dominant, varied largely depending on the initial conditions of the different vegetation types, and on the fire regimes. For example, pines disappeared as soon as two subsequent fires occur within about 12 years (i.e. pine maturity age).
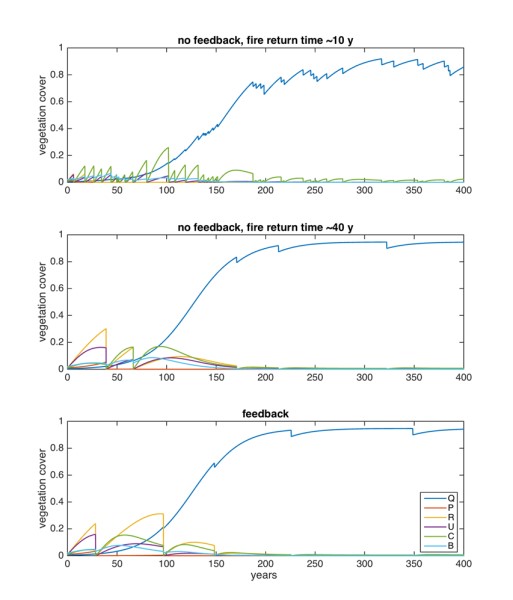
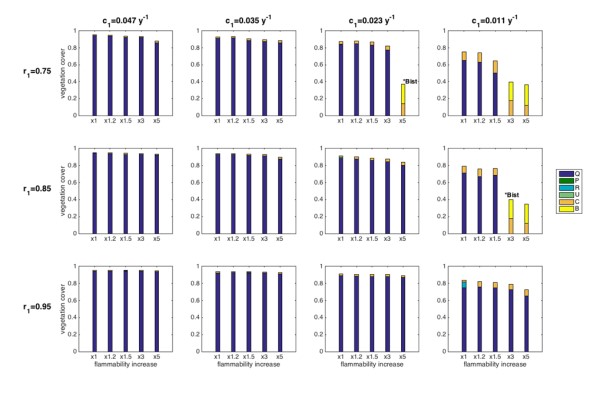
To simulate what would be the effect of drought on the species composition a system subject to recurrent fires, we implemented drought effects on:i) oak reproduction and establishment, via lowering of the colonization ability parameter; ii) via lowering the oak resprouting ability parameter; iii) via fire recurrence, either directly by increasing the average return time (no feedback case, not shown), or indirectly by increasing the probability of fire in the presence of the different plant types (feedback case) (Figure 4). For both fire dynamics, we observed that reduced resprouting ability and lower oak colonization rates could lead to a very different community, namely composed of Cistus and Brachypodium, with more than 50% of the available space left empty. When including the fire feedback, an increased flammability of all species was necessary to obtain this different state (Figure 4).
In some cases, the existence of these communities depended on the initial conditions: if the ecosystem was initially mainly composed of oaks, the forest remained and fires were relatively infrequent (~every 40 years), while if the initial abundance of seeders and/or Brachypodium were high, the system stayed locked in the shrubland/grassland state (“degraded”), with more frequent fires (~every 15 years). This is a so-called stochastic bistability (see Figure 4 where the panels displaying bistability are marked with an asterisk). This bistability implies that catastrophic shifts may occur in the system under drier conditions than the current ones: a shrubland may then not evolve naturally into an oak forest anymore, as expected under the current climatic conditions, but it would stay locked in its (degraded) state. If the drought further increased the flammability, or decreased the oak resprouting ability, the system inevitably (i.e. for any initial conditions) developed into the degraded state, and it never reached the forest state. The no-feedback case (not shown) was similar, but it did not display any bistability, thus indicating that bistability was a consequence of the fire feedback itself. Also, for the system to evolve into a degraded state, increasing the fire frequency was not a strictly necessary condition: when oak recruitment and resprouting were limited, the degraded state was reached in conditions where fires were relatively rare (e.g. every 40 years). Finally, the degraded state could also be composed of Cistus alone or with a minor (<10%) fraction of oaks. This suggested that when the fire feedback was included, the grass survived because of it could withstand the highly recurrent fire that were generated by its high flammability thanks to its resprouting ability.
Conclusion
This second fire modeling effort suggests again that, under the current conditions, the successional process in the Mediterranean area generally leads to a dominance of the oaks. Also, pine forests disappear in all cases as a consequence of their highest vulnerability to fires (as in Vasques et al. [46] and [88]). However, this study also shows that drier conditions, expected as a consequence of climate change, can diverge the succession, leading to an open shrubland instead of the expected late-successional oak forests. This is due to drought reducing the resprouting capacity, the growth and seedling establishment of oaks, in combination with an enhanced flammability, which is expected based on current observations (see e.g. »Structural and functional changes associated with regime shifts and »Restoration potential for preventing and reversing regime shifts). Within a certain range of drought intensity, and if the feedback between species community composition and fire is considered, shrubland is effectively an alternative (stochastically) stable state to the closed forests, implying that catastrophic shifts between these states are possible if drier conditions occur. The degraded shrubland is mainly composed of Cistus and Brachipodium. The dominance of rockrose (Cistus spp.) over the other shrub seeders is expected, as a consequence of the low regeneration rate of Ulex parviflorus and of the difference in plant longevity, combined with their flammability [89].
Note: For full references to papers quoted in this article see