| Contributing Authors: | Sonia Kéfi, Florian Schneider, Angeles G. Mayor, Alain Danet, Marina Rillo, Simon Benateau, Jacob Keizer, Ana Vasques, Susana Bautista, Paco Rodriguez, Alejandro Valdecantos, Jaime Baeza, Ramón Vallejo, Max Rietkerk, Mara Baudena, Mart Verwijmeren, Koen Siteur, Rubén Díaz-Sierra |
| Editor: | Jane Brandt |
| Source document: | S. Kéfi, A. Vasques, F. Schneider, M. Rietkerk, A.G. Mayor, M. Verwijmeren, R. Diaz-Sierra and M. Baudena. 2016. Response of Mediterranean drylands to increasing pressures. CASCADE Project Deliverable 6.1, 54 pp |
Grazing is one of the main causes of desertification of drylands worldwide [1,13]. Understanding how and why different types of grazing and different levels of grazing pressures affect dryland resilience is therefore of great importance to inform dryland management and help preventing desertification. One of the objectives of CASCADE was to develop and analyze models that help us investigate the role of grazing in dryland dynamics and resilience, by improving the realism with which grazing is classically incorporated in dryland vegetation models.
In a number of dryland models [8,11], plant mortality is taken as a proxy for grazing pressure. Grazing, in these models, simply increases plant mortality. Moreover grazing is typically considered to be homogeneous in space, meaning that it is affecting all individuals equally [7,8,11,14].
However, grazing can have a strong spatial component [15–18]. In arid shrublands that have historically been exposed to grazing by large herbivores, most plant species have developed mechanical defenses against large herbivores, such as fast regrowth from the root stock, indigestible tissue, the development of prostrate growth forms and the evolution of spines and thorns [19,20]. Thus, coinciding with the effects of abiotic facilitation, i.e. the amelioration of the local environmental conditions through shading or water retention by the presence of vegetation [21], the canopy of plants with defenses against large herbivores also provides refugia from large herbivores to neighboring plants [21–24].
This mechanism leads to low individual mortality in places where local plant cover is high. We refer to this feature of a spatially constrained negative density dependence of mortality as ‘associational resistance’ (see [21] for a discussion on the term). Vice versa, a low local plant cover will increase the risk of dying due to grazing for the remaining vegetation [21]. As a consequence, if the overall benefit of grazing protection outweighs the cost of competition for limiting resources, plants coincidentally team up with other plants [24], which contributes to the formation of patch pattern at the landscape scale [21]. In such case, the highest risk of being grazed is borne by plants that grow isolated from others, whereas plants that are growing at the border or in the center of a patch are less vulnerable to grazing or even entirely unaffected because they benefit from associational resistance [21,23].
We introduced ‘associational resistance’ in a spatially explicit dryland vegetation model, i.e. the mutual reduction of grazing impact by sharing defensive traits [25]. This dryland vegetation model was used to investigate the interactive effects of grazing (now a spatially heterogeneous pressure) and aridity on the spatial vegetation patterns and on ecosystem resilience. The model was inspired by the Randi forest CASCADE site in Cyprus, but can be generally applied to grazed patchy drylands.
Model description
The model is an interaction particle system that describes the landscape as a grid of cells, each of which can be in one of three states:
- ‘vegetated’ cells are occupied by a plant (black cells in Figure 1c);
- ‘empty’ cells do not contain adult plants but are suitable for seeds to germinate and establish (grey cells in Figure 1c);
- ‘degraded’ cells represent bare ground which has been eroded, lacks organic matter, is characterized by bad water retention, and therefore cannot be colonized by arriving seeds (white cells in Figure 1c).
Transitions between cell states are only possible between vegetated and empty (by the processes of plant ‘death’ and ‘recolonization’) and between empty and degraded (by ‘degradation’ and ‘regeneration’). In biological terms, a degraded site needs to be enriched first, before a plant can establish on it. Conversely, when a plant dies, it leaves the spot empty but still enriched, until it becomes degraded by erosion.

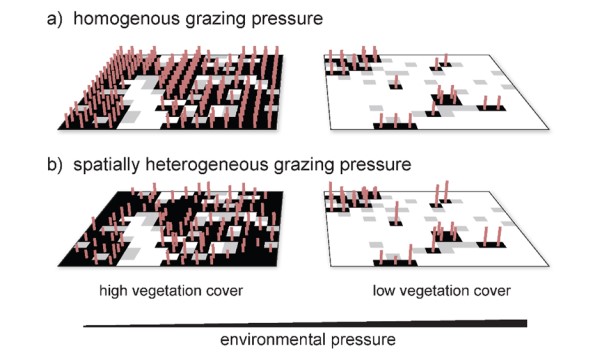
Associational resistance was introduced in the model as follows: we assume that plants in the center of a patch therefore bear a lower risk of being grazed by herbivores than plants growing isolated from others (Figure 2). By including this mechanism, grazing becomes a spatially heterogeneous pressure. The probabilities for these transitions to occur on any cell currently at state might be constants or functions of the global vegetation cover or of the local vegetation cover in the neighborhood of the focal cell (equations can be found in [25]).
Results
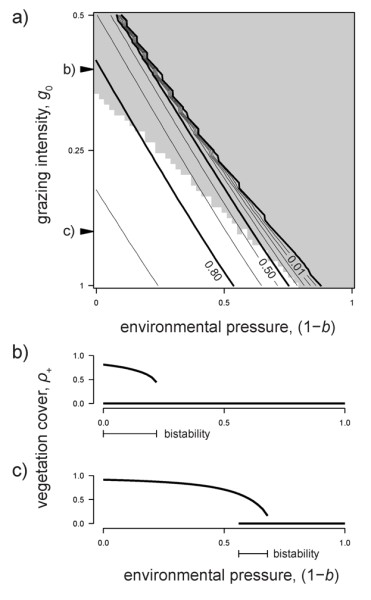
We found that spatially heterogeneous grazing altered ecosystem stability. Indeed, with associational resistance, plants' risk of being eaten depended on the local configuration. Therefore, grazing impact was minimized at high cover but became self-enhancing in more fragmented landscapes. This feedback on plant mortality disrupted patch growth and put even apparently 'healthy' drylands under high risk of catastrophic shifts (Figure 3). More specifically, the range of pressure levels at which both desert and vegetated landscapes were simultaneously stable (i.e. the so-called bistability area) increased as spatially explicit grazing became more intense. This is consistent with previous modelling studies of spatially homogeneous grazing pressure [8,11,26]. Second, under high grazing pressure, state transitions from a vegetated to a bare landscape were more sudden and unexpected and the ecosystem shift to desert occurred at higher vegetation cover and connectivity, i.e. in apparently ‘healthy’ landscapes.
Conclusion
Our results indicate that when ignoring the interfering feedback mechanisms caused by the spatially explicit pressure (here grazing), we might over-estimate ecosystem resilience and impede the success of sustainable management practices.
Note: For full references to papers quoted in this article see